Hei,
Mitt navn er Maria Hole og jeg jobber med å få ferdig min bachelor i molekylærbiologi dette semesteret. Jeg har lenge hatt lyst til å gjøre mer praktisk arbeid i studiene mine, så jeg benyttet derfor muligheten til å være en del av et prosjekt hos professor Jon Vidar Helvik som omhandler lysoppfattelse og melatoninsyntese i Atlanterhavslaks. Mitt prosjekt går ut på å klone genene til enzymet AANAT som er hastighetsbegrensende i syntesen av melatonin. Klonene av disse brukes videre for å lage prober som benyttes i en metode kalt in situ hybridisering som benyttes for å se hvor disse genene uttrykkes i laksehjernen. Resultatene av mitt prosjekt skal brukes videre for å se om disse genene uttrykkes på samme plass som ulike opsiner (lysoppfattende celler) for å gi en større forståelse av hvordan lys påvirker laks på ulike stadier i livet.
Bildene under viser noe av det jeg har gjort i dette prosjektet
Kloning av aanatgener
Hele kloningsprosessen er ganske tidkrevende og komplisert, siden det er mange metoder som må gjennomføres. I tillegg må man hele tiden sjekke om metoden som er gjennomført faktisk har fungert før man kan gå videre i prosessen.

Pipettering: Alle metodene jeg har vært innom har innebært mye pipettering av ulike løsninger. Dette er fordi det er svært små volum som skal måles opp, gjerne ned til 1μL

Gel elektroforese: Denne metoden brukes for å verifisere at prøvene mine innholdt riktig DNA. Det gjøres ved at de føres gjennom en porøs gel ved hjelp av elektrisitet (DNA er negativt ladet, så beveger seg mot pluss-polen). En DNA ladder med innhold med kjente størrelser benyttes for å gjenkjenne størrelsen til DNAet.
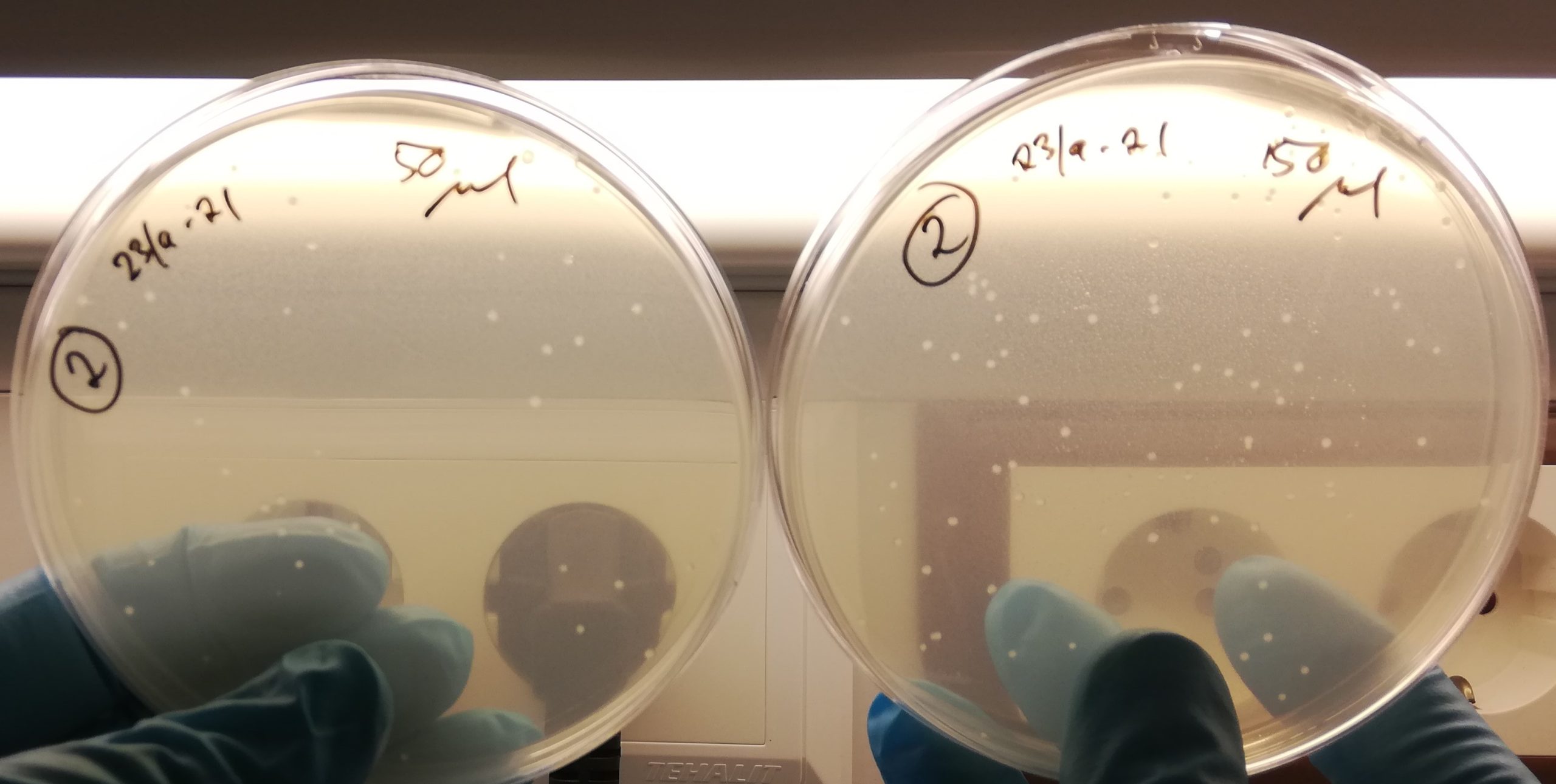
Bakteriekolonier for kloning av aanat: De hvite prikkene er bakteriekolonier. aanat gensekvensen er plassert i et plasmid som har fått vokse i bakteriekolonier i optimale forhold. LB-mediet de ligger i inneholder ampencilin som dreper alle bakterier som ikke er antibiotikaresistente, noe plasmidet jeg brukte er.
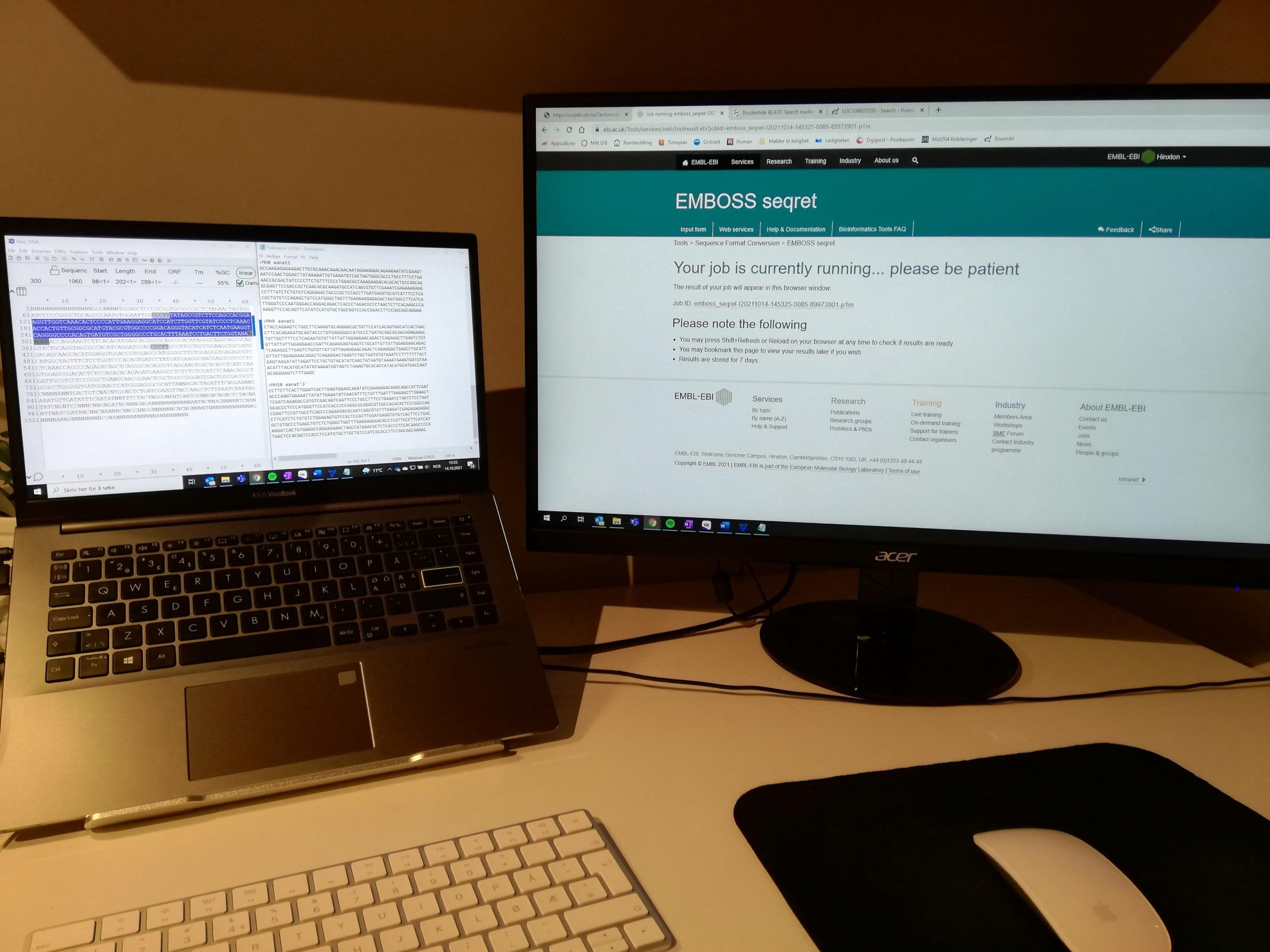
Sekvensbehandling: Også mye av arbeidet foregår på PC. Her har jeg fått sekvensen på de genene jeg har klonet fra sekvenslaben. Arbeidet her går ut på å sjekke at genet jeg har klonet faktisk er aanat og tilrettelegging for designing av primere som må bestilles før proben skal lages.
In situ hybridisering
For å lokalisere aanat blir en probe brukt som guide til visualisering av genuttrykket. Proben er komplimentær til aanat-sekvensen som befinner seg inni cellene, og vil derfor feste (hybridiseres) til disse sekvensene. Proben har en markør som benyttet for visualisering, og dermed kan man se hvor i hjernen proben befinner seg. Denne må syntetiseres før man kan sette i gang med in situ hybridisering, som gjerne tar to dager. Prosessen innebærer mange steg der vevet må forberedes for at proben skal kunne trenge inn i cellene, i tillegg til mye inkubasjonstid for hvert av stegene. Hele in situ hybridiseringsprotokollen tar 3 dager å gjennomføre.

Snitting av laksehjerner: For å lokalisere genuttrykk i hjernen på laks, må hjernen snittes i små snitter på 10μm. I dette kammeret er det -20˚C, siden mediet laksehodene er montert i da er i fast form.

Løsninger til in situ hybridisering Behandling av snittene med mange ulike løsninger, der det å lage til disse løsningene også er en del av prosessen
In situ hybridisering Her står snittene i en kuvette med en av løsningene brukt i in situ protokollen. Det er ulikt hvor lenge de skal stå i hver løsning, noen må stå slik i en halvtime.

Inkubasjon av behandlede snitt. Etter første dag skal snittene inkuberes over natten i et lufttett kammer, etter at proben er tilsatt.

Ferdig produkt. Her er alle snittene etter at in situ hybridisering er gjennomført.
Hvordan ser man at in situ hybridiseringen har funket, og kan vi stole på det?
Der aanat uttrykkes, vil man gjennom mikroskop kunne se lilla markeringer på snittene. Hjernestrukturene på snittene er mulig å se ved hjelp av mikroskop, slik at man kan se hvor markeringen tilsier at genet uttrykkes. Alt i alt brukte jeg tre ulike prober siden vi ville undersøke tre ulike aanat genarianter. For hver antisense-probe (komplimentær til aanat-sekvensen) ble samme prosess gjennomført med en sense-probe (lik sekvens som aanat-sekvensen) som ikke fester seg til aanat-sekvensen i hjernecellene. Dette fungerer som en kontroll som sikrer at det er antisenseproben som forårsaker markeringen, og dermed vet vi at genet uttrykkes der markeringen er.
Mine tanker og erfaringer om prosjektet og faget
Jeg syns det har vært veldig gøy å drive med dette fordi jeg har fått muligheten til å jobbe en del på egenhånd. Dette har gjort det mulig for meg å styre dagene litt selv, samtidig som jeg alltid har hatt folk å spørre om hjelp. Det aller gøyeste har vært å faktisk forstå hvorfor jeg gjør de metodene jeg gjør og hva de ulike stegene i protokollen gjør med produktet mitt. Dette har gjort det lettere å stille vite hvilke spørsmål jeg skal stille for å forstå den store helheten av arbeidet mitt.
Er det noe jeg har lært i dette faget, så er det hvor lite tidsoptimistisk man kan være når man er på lab. Stort sett så tar alt mye lengre tid enn man tror, spesielt når man har lite erfaring med de metodene man bruker. Det er alltid noen faktorer som kan sørge for at alt tar veldig mye mer tid, enten at man har glemt å hente en løsning fra fryseren og må vente på at den tiner, eller at bestillinger har lang leveringstid på grunn av Brexit (som ga meg en lab-pause på to uker). Man kan planlegge og forberede seg for å forhindre forsinkelser, men man må alltid være forbered på at ting ikke går den veien man ønsker. Dette har virkelig lært meg viktigheten av god planlegging og ikke minst god forberedelse slik at man vet hva som skal gjøres til enhver tid i løpet av en dag.
Det som har vært spennende med mitt prosjekt er at jeg kan ikke alltid vite om det jeg gjør har fungert, noe som danner en viss uvitenhet til sluttproduktet. Jeg har hatt muligheten til å verifisere at jeg har det produktet jeg skal ha, eller at genet mitt ble satt inn i plasmidet og dermed klonet, men det er først etter at in situ resultatene kom at jeg føler jeg har mestret projektet og at alt det jeg har jobbet med har fungert. Resultatene blir veldig visuelle og selvforklarende, i motsetning til et eppendorfrør med 1 mL gjennomsiktig væske som jeg jo vet at inneholder det det skal inneholde. Derfor er jeg veldig takknemlig for at jeg har fått muligheten til å gjennomføre hele prosjektet, til tross for mye forsinkelser. Jeg vil takke mine veiledere, professor Jon Vidar Helvik, forsker Mariann Eilertsen og Ph.D. stipendiat Christine Horne for faglig veiledning og god hjelp med arbeidet mitt, og ikke minst takk til teknikker Rita Karlsen for god opplæring og veiledning på laben og i alle metodene jeg har brukt.